04.04.2018
A clinical overview of the autonomic nervous system, the supply to the gut and mind–body pathways
LOUIS GIFFORD AND MICK THACKER
Introduction
The autonomic nervous system (ANS) is an ‘output’, ‘effector’ or ‘motor’ system (see Gifford 1998b)—i.e. it responds to the demands of the sensory systems and the central nervous system (CNS) by producing an effect on the tissues it supplies. There are no sympathetic or parasympathetic sensory (afferent) fibres as such, although a great many visceral sensory fibres do travel in nerves like the vagus and splanchnic, which are commonly described in parasympathetic and sympathetic sections of anatomy texts and often referred to as ‘sympathetic afferents’ (see discussion of visceral afferents below).
Since the ANS is an output system, its role in pain states has to be secondary to its effects on sensory systems (see following chapters).
ANS fibres create their ‘effects’ via electrochemical stimulation of smooth muscles or glands. Their secretions also produce direct chemical effects on the tissues they innervate and thus change their chemical characteristics. Any chemical or physiological tissue changes may in turn be fed back to the CNS via stimulation of sensory afferent pathways. This is important because ANS activity has been implicated not only in producing and modulating nociception and pain, but also as playing a role in the expression and conscious awareness of emotions (see Meyers 1986). Recent studies show, for example, that different emotions (anger, fear, disgust, sadness, happiness, surprise) can be distinguished to some extent on the basis of different autonomic nervous system responses (like skin temperature and heart rate) (LeDoux 1998, p 292). We all know that strong emotions are expressed as bodily sensations and that ‘gut feelings’ play crucial roles in our emotional experiences and can have a strong impact on our decision making processes (Damasio 1995, LeDoux 1998, Damasio 2000).
In their detailed overview of the ANS, Janig and Habler (1999) go out of their way to debunk the traditional descriptions of the ANS and the view that it is ‘all or nothing’ and general in its mode of action. In parallel with the neuroendocrine system, the ANS regulates target organs in order to maintain the homeostasis of the body. Simply, it adapts, adjusts and co-ordinates appropriate systems so as to produce the required physiological conditions for life, which includes, any required physical activity. In this way it is involved in producing and co-ordinating the necessary physiological responses for activities like moving, resting, sleeping, feeding and digestion, sex, pregnancy and nurturing, growth and repair and all extreme responses relating to threatening and stressful situations. This includes responding to tissue injury and to pain.

Adapted from: Janig W, Habler HJ 1999 Organization of the autonomic nervous system: structure and function. In: Appenzeller O (ed) Handbook of Clinical Neurology Vol 74 (30): The Autonomic Nervous System Part 1: Normal Functions. Elsevier, Amsterdam
ANS activities can be viewed as being governed by the commands it receives from the modules of the CNS that ‘scrutinise and appraise’ the sensory and incoming information received (Gifford 1998). Thus, environmental inputs from sense organs, electrochemical sensory inputs from body tissues and organs, inputs derived from the immune system and inputs that derive from the thinking, analysing and planning centres of the brain, all serve to create specific ANS driving stimuli. Thus, at least in part, the ANS is subservient to the wishes and desires created in our mind. If you want to get up out of the chair and go for a walk—the ANS has to organise the physiological backup and supply networks that support the action demanded.
Figure 1.1, taken from Janig and Habler (1999) and modified slightly, usefully illustrates where the ANS occupancy of the nervous/homeostatic systems sits. The model, which is quite like the Mature Organism Model (Gifford 1998) in that it is ‘circular’—having input, processing and output elements—nicely illustrates the integration of all elements of the body including the brain and its higher centres.
The following example may help you/the reader appreciate the role of the ANS and the importance of models like these to clinical reasoning.
In creation of hunger, ‘body organs’ relay information to the CNS about the levels of glucose in the blood. Information may be via neural or humoral/ endocrine systems and messengers. From the neural pathway perspective, it is known that special chemoreceptive sensory nerve fibre endings that respond to blood glucose levels exist in the pancreas and gastrointestinal tract as well as in the blood vessels of the third ventricle of the brain adjacent to the hypothalamus. The ‘sampled’ information is then relayed to specific regions of the CNS that scrutinise and deal with ‘actions’ relating to hunger and energy supply. The message is simply—‘energy levels low, please restore.’ The CNS regions involved in scrutinising and mounting a coordinated response are thought to reside in the hypothalamus, in particular, the paraventricular and ventromedial nuclei and the lateral hypothalamic areas (Westmoreland et al 1994). ‘Output’ from these areas may be directed to the limbic and neocortical areas, creating the sensation of hunger, which in turn drives food gathering behaviour via the musculoskeletal system and thence to the ‘external world’ as noted in Figure 1.1.
At the same time:
- Autonomic output activates relevant regions of the alimentary tract in order to make ready for food intake and digestion.
- Autonomic activity will also stimulate the pancreas and adrenal glands, both of which are important in the hormonal regulation of glucose and energy metabolism.
- Autonomic activity will co-ordinate the cardiovascular system so that blood supply demands may be met from the musculoskeletal system whose activities are needed to gather food, prepare, eat, and metabolise it.
As the process of food gathering and feeding proceed so the input, scrutinising, and output systems change their activities in response to the changing situations.
It is often said that the sympathetic system ‘kicks into action during emergencies’. Robert Salposky (1994) adds the (clinically) important caveat ‘or what you think are emergencies’—that underlines the importance of personal assessment of the situation we find ourselves in. Thus, how the sympathetic system responds is very much influenced by the emotional, cognitive and conscious brain—‘our’ interpretation of the situation we are in is hugely responsible for its activities. If you believe the pain you have signifies a serious illness or condition you will have a far different ‘sympathetic’ reaction than if you dismiss the pain and pass it off as trivial. If as a patient you think that what the therapist is doing to you makes sense and feels right you will have quite a different response from someone who might be feeling unsure or even slightly anxious about what is being done to them (see also Chapter 4). It is probable that a clinician’s most powerful effect on a patient’s sympathetic system activity is produced via the atmosphere of the therapeutic interaction. Importantly, this relates more to what the patient actually feels and interprets, than to what the clinician thinks they should feel or should have interpreted. The effect of a productive ‘therapeutic alliance’ on the activities of systems like the ANS should never be underestimated (see the Placebo section in Topical Issues in Pain 4.)
Autonomic anatomy—from spinal cord out
As discussed, the sympathetic and parasympathetic systems are efferent— just like the somatic motor system they have an ‘effect’ on the tissues that they innervate. Simply, impulses radiate from the central nervous system to target tissues in the body. While activity in somatic motor efferent neurones cause striated muscles to contract, those in the autonomic nervous system have a wide range of targets and effects. For example, via the sympathetic innervation of smooth muscles in blood vessel walls sympathetic activity can alter vessel lumen size and hence alter blood flow; it can cause secretory glands to be activated or inactivated; and, via secretion of chemicals from vesicles in its terminals, it can influence the chemical environment, the health and the healing of the tissues innervated (see Chapters 2 and 3). As well as having the capacity to have quite massive effects, it also seems that the autonomic nervous system is capable of acting in a highly specific and localised way.
The sympathetic system
This section is intended to provide the reader with an overview of a poorly understood system. Review of classic anatomical texts and overviews by current leaders in the field of autonomic nervous system anatomy, physiology and function reveals quite marked discrepancies and inconsistencies. The reader would be well advised to read some of the papers and texts cited at the end of this chapter for fuller accounts
Adapted from: Harati Y, Machkhas H 1997 Spinal cord and peripheral nervous system. In: Low PA (ed) Clinical Autonomic Disorders 2nd edn. Lippincott-Raven, Philadelphia
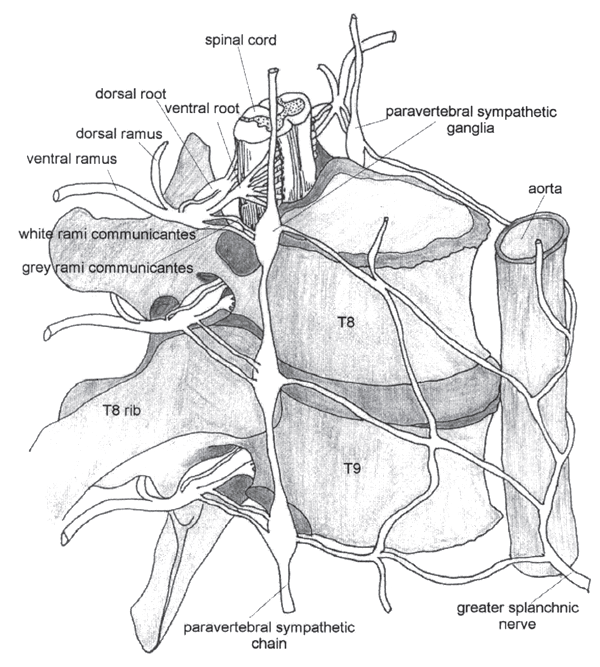
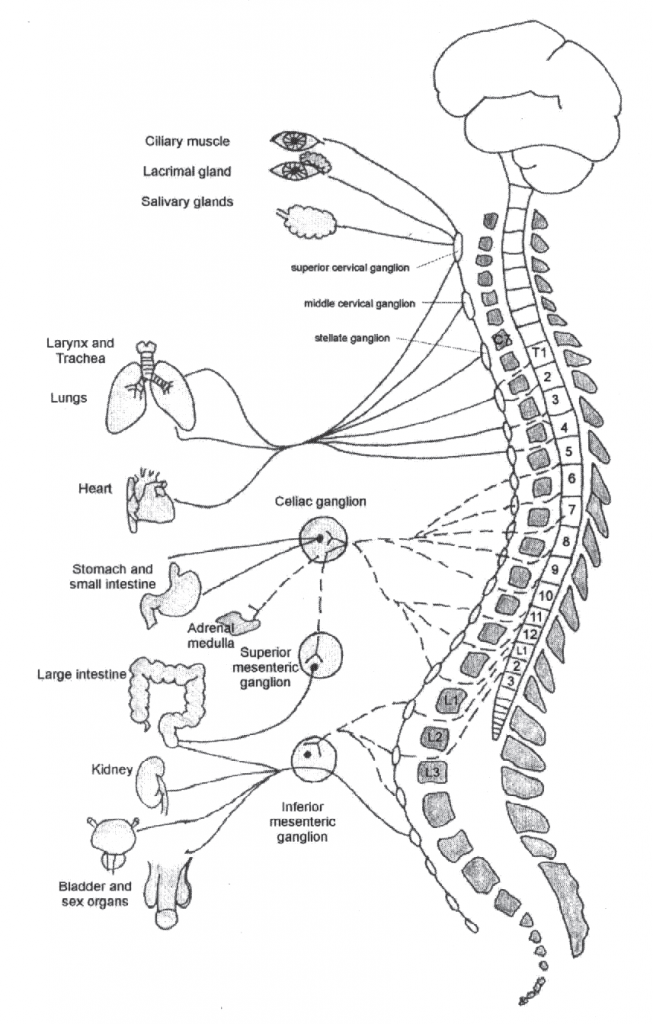
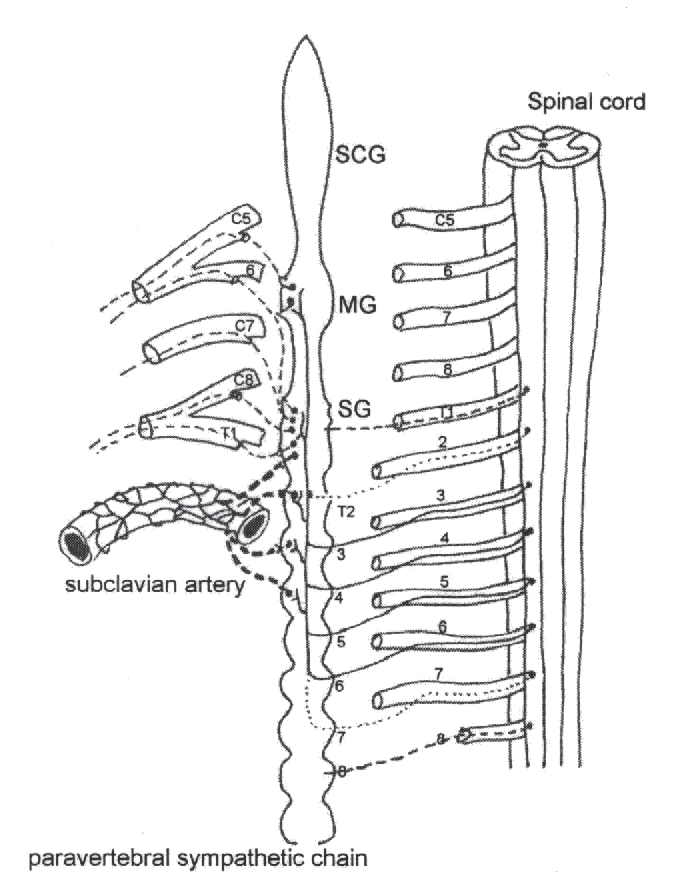
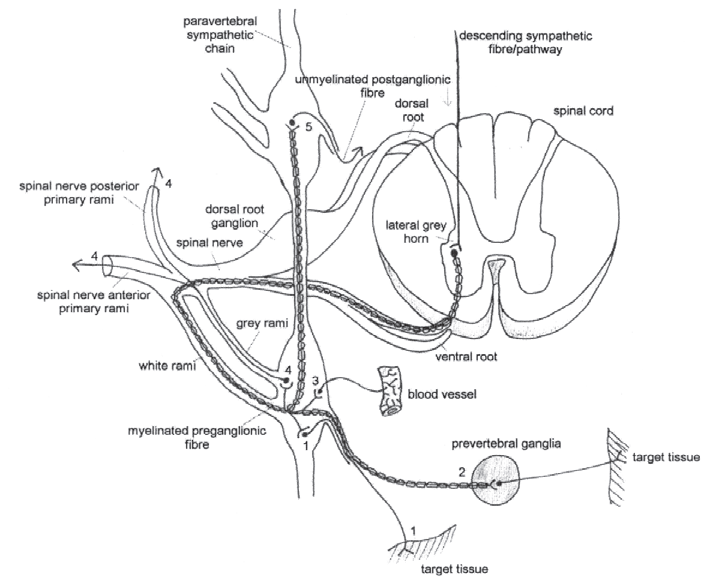
The best known parts of the sympathetic system are the two sympathetic chains or trunks that extend from the base of the skull to the coccyx. These two ‘paravertebral’ sympathetic chains lie on either side of the vertebral column (Figs 1.2, 1.3, 1.4 & 1.5) but may come together and fuse in the sacral region to form the ganglion ‘impar’. Each chain has about 22 or 23 paravertebral ganglia which contain nerve axons and the cell bodies of postganglionic neurones. A ganglion is a swelling of a nerve due to the large numbers of cell bodies it contains. Sympathetic ganglia can be thought of as communication boxes or relay stations where information in the form of impulses may be modulated and passed on, even prevented from passing on, or simply allowed to pass straight through unchecked. Many of the paravertebral ganglia relate to the cord segment from which they derive (Fig. 1.3). However, there are fewer in number than there are segments in the spine as some of the ganglia are fused. In the cervical region there are normally three ganglia—the most rostral being the superior cervical ganglion, below this is the middle cervical, and lowest of all, the stellate ganglia (Figs 1.3, 1.4). The stellate ganglion consists of the fused inferior cervical ganglion with the first thoracic ganglion. There are usually four ganglia in the lumbar part of the chain and four or five in the sacral region.
Adapted from: Harati Y, Machkhas H 1997 Spinal cord and peripheral nervous system. In: Low PA (ed) Clinical Autonomic Disorders 2nd edn. Lippincott-Raven, Philadelphia
Adapted from Grieve GP 1994 The autonomic nervous system in vertebral pain syndromes, Fig. 20.4. In: Boyling JD, Palastanga N (eds) Grieve’s Modern Manual Therapy 2nd edn. Churchill Livingstone, Edinburgh
Adapted from Grieve GP 1994 The autonomic nervous system in vertebral pain syndromes, Fig. 20.5. In: Boyling JD, Palastanga N (eds) Grieve’s Modern Manual Therapy 2nd edn. Churchill Livingstone, Edinburgh
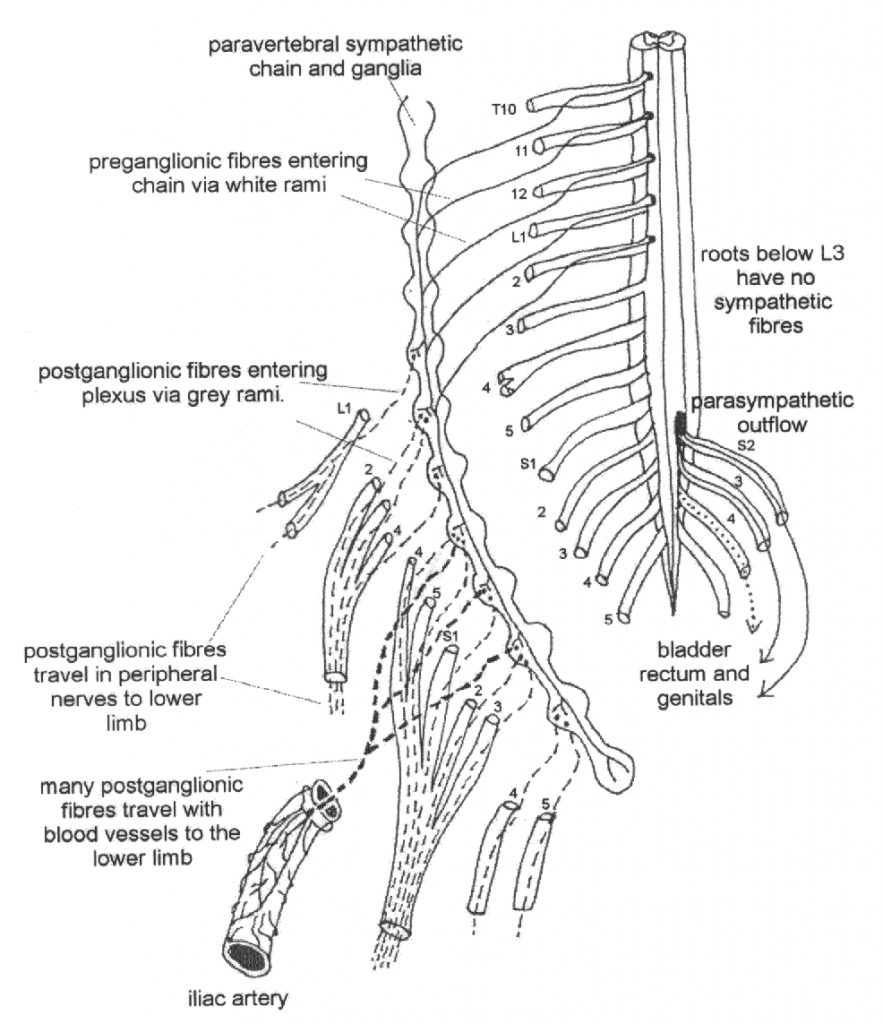
The spinal cord only connects to the sympathetic chain via the 14 spinal nerves that exit from the intervertebral foramen between T1 and L3 (i.e. via roots T1–L2) (Figs 1.3, 1.4, 1.5). Hence its ‘outflow’ is said to be ‘thoracolumbar’. This means that all the peripheral sympathetic nerve pathways to the target tissues and organs of the body have their origins in the spinal cord segments of T1 to L3 (but see Grieve 1994, p 297).
Although it has segmental origins, the SNS, to quite a marked extent, actually defies the traditional description of a segmental system (see below). Thus, sympathetic supply to the head and neck has its origins from the nerve roots of T1–T5 (mostly upper 2 or 3); the upper limb from roots T2–T6 (but possibly as far as T7 or T8) (Fig. 1.4); the thorax from roots T3–T6; the abdomen from roots T7–T11 and the lower limbs from roots T10–L2 or L3 (Fig. 1.5).
This supply may be an important consideration when investigating the consequences of thoracic and high lumbar nerve trunk and nerve root injuries or irritations. What can be applauded is the neat fact that these vital control supply lines exit and have their origins from parts of the spinal column that are relatively rarely injured and well protected.
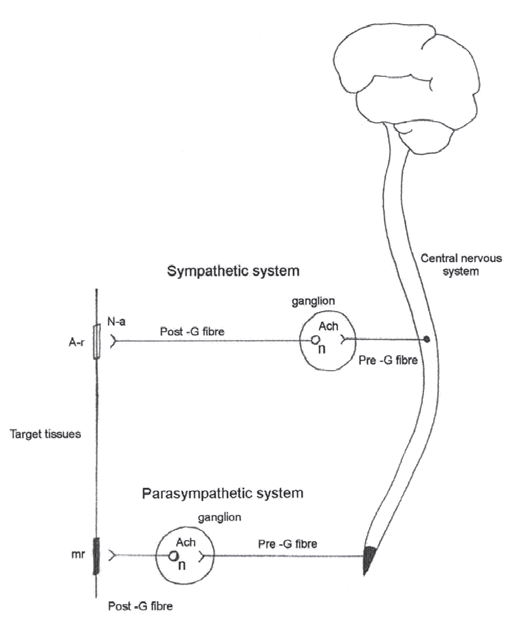
The sympathetic supply involves a two neurone pathway from the spinal cord to the target tissues of the periphery (Fig. 1.6). These neurones are termed preganglionic and postganglionic respectively.
There is some evidence that the sympathetic ganglia contain interneurones, termed ‘SIF’ (small intensely fluorescent) cells that may serve modulatory functions, i.e. they aid in the integration or processing of incoming and outgoing information (see Grieve 1994, p 299).
The preganglionic neurones of the sympathetic system have their cell bodies in the lateral grey horn (Fig. 1.7), (also known as the intermediate zone, or interomediolateral cell column) of the spinal cord. As already noted, the cell bodies of these neurones are only found in segments T1 to L3 (Westmoreland et al 1994) of the spinal cord. Its segmental origins are worth noting since the sympathetic system innervates just about every tissue of the body via these spinal segments.
Myelinated preganglionic fibres pass away from the spinal cord via the ventral root and into the spinal nerve for a short distance, then via the white rami communicantes they join the sympathetic trunk.
Having reached the sympathetic trunk, the preganglionic sympathetic fibres may do several things (see Fig. 1.7). Some synapse with postganglionic cells in the paravertebral ganglion that lies at the same level from which they exit the vertebral column (nos 1, 3 & 4 in Fig. 1.7). Others pass through their segmental ganglion and run up or down to more distant paravertebral ganglia where they terminate and synapse with postganglionic fibres (no. 5 in Fig. 1.7). Note how this defies the idea of ‘segmentalism’. It is commonly stated that there is a pre to postganglionic neurone divergence ratio of anything from 1:1 to 1:196 (see Grieve 1994, Williams et al 1995). In other words one preganglionic fibre may terminate with only one postganglionic fibre, or may have a massive terminal aborisation connecting to a great many. The reality is that there is anatomical convergence (many preganglionic fibres going to one postganglionic fibre), as well as anatomical divergence (the opposite). Importantly, anatomical convergence/divergence does not mean that all the connections are necessarily always functional. Just as elsewhere in the nervous system, there is a huge potential for a spread of effect or a focus of effect that is dependent on continuously fluctuating inhibitory and excitatory modulatory controls. This attribute allows for both expansion and exquisite refinement of autonomic outputs and argues against the blind acceptance of the generalised fight and flight-related function of this system. The possible modulatory role of ganglionic inter-neurones as well as branches from returning visceral sensory afferents (see below) may be important here.
What we do know is that the SNS tends to be a functionally ‘divergent’ system—where small numbers of preganglionic fibres have the potential to influence larger numbers of postganglionic fibres—hence the potential for a single ‘action’ message producing far reaching effects on target organs and tissues. The fact that there may be provision for control of this divergence helps explain a degree of specificity, too.
Adapted from: Harati Y, Machkhas H 1997 Spinal cord and peripheral nervous system. In: Low PA (ed) Clinical Autonomic Disorders 2nd edn. Lippincott-Raven, Philadelphia
In a similar way to that described in the CNS, the potential for excessive reactivity and spread of effect due to maladaptively altered modulatory controls, or even loss of control, having far reaching effects on sympathetic activity seems a reasonable possibility. Clinically this may provide a possible explanation for reports of localised patches of skin colour change, or localised sweating or swelling disturbances that spread to larger areas over time.
Many preganglionic fibres pass right through the sympathetic chain and paravertebral ganglia without synapsing on postganglionic fibre cells at all (no. 2 in Fig. 1.7). These continue out of the sympathetic chains in the splanchnic nerves to reach remote ganglia situated nearer their target organs. These more distant ganglia are often called ‘prevertebral’ ganglia (Figs 1.3, 1.7). Well known prevertebral ganglia supplied via the thoracic splanchnic nerves are the celiac, superior and inferior mesenteric ganglia. These ganglia are situated in the abdomen anterior to the abdominal aorta close to the origin of the celiac and mesenteric arteries. Via postganglionic fibres they form the celiac plexus that innervates abdominal, pelvic and perineal organs. While the thoracic splanchnic nerves derive from branches arising from the lower eight thoracic paravertebral ganglia, the lumbar splanchnic nerves derive from branches arising from the upper three lumbar paravertebral ganglia.
One special group of preganglionic neurones pass directly to their target tissue (see Fig. 1.3). These innervate the adrenal medulla, seemingly omitting the ‘postganglionic’ component of the pathway. In fact, the adrenal medulla resembles nervous tissue more than it does a typical endocrine gland. Its cells behave like neurones and derive embryologically from postganglionic sympathetic cells. These adrenal medulla cells secrete adrenaline (epinephrine) and noradrenaline (norepinephrine) into the circulation in response to SNS activity that derives from situations that are exciting or stressful, or that are anticipated to be exciting or stressful. This is an excellent example of how mind influences body and can easily be translated to the clinical encounter!
Postganglionic fibres have their cell bodies in the autonomic ganglia described above. Their unmyelinated axons radiate to the target tissues. Those postganglionic fibres that have their origins in the ganglia of the paravertebral sympathetic chain follow three main courses:
- Pass back into the spinal somatic nerve trunk via grey rami communicantes (no. 4 in Fig. 1.7). Some apparently may pass back in via the white rami communicantes, too (Janig & Habler 1999). From there, the fibres travel in the peripheral nerves to supply the target tissues, for example, the blood vessels (vasomotor), and sweat glands (sudomotor) in the territories of the nerves they accompany. Thus, postganglionic axons from the cervical ganglia innervate upper extremity tissues by entering the cervical spinal nerves of the brachial plexus deriving from C4–C8, and following the peripheral nerve trunks (see Fig. 1.4). Grey rami communicantes in the neck may pierce and travel through the longus capitis or the scalenus anterior muscles—a fact that may be an important consideration in whiplash injury (see Thacker 1998). Clearly, if these grey rami are injured it may mean the loss of sympathetic supply and control to blood vessels and tissues of the arm.
- Pass to blood vessels in the neighbourhood of the sympathetic trunk (no. 3 in Figs 1.7 and 1.2, 1.4, & 1.5) and supply these or travel along with them to reach more distal targets. For example, axons from the superior cervical ganglion innervate the pupil of the eye and provide sweat gland innervation of the face by following the course of branches of the internal and external carotid arteries to get there.
- Pass to visceral organs. For example, postganglionic fibres whose origins are in the lower cervical and upper thoracic ganglia innervate the heart via the cardiac plexus to produce cardiac stimulation, or reach the tracheobrachial tree via the pulmonary plexus to control bronchodilation (we need more efficient lungs when we are about to perform physically, e.g. as in competition or combat).
Postganglionic fibres may pass to higher or lower levels within the sympathetic trunk before leaving via the above routes. For example, although we traditionally describe the efferent supply to the arm as arising from C4–C8 it may be from as low as T7 or T8 if we trace the sympathetic supply back to the roots and spinal cord.
Clinical implications
Some clinically related points:
- It is important to emphasise that all peripheral nerves contain sympathetic post ganglionic fibres and that they enter the nerves via grey rami communicantes.
This means that those somatic nerves that derive from nerve roots above T1 (including many cranial nerves) and from roots of L3 and below, still receive their supply via grey rami communicantes but will have no white rami communicantes (Figs 1.4 & 1.5). Think about it and you realise that all the cervical nerve roots and those roots below L3 will have no sympathetic neurones. The sympathetic nerve fibres join the spinal nerves outside the intervertebral foramen. This may well be an important clinical/diagnostic consideration. For example, it may explain the common clinical finding that classic acute and sub-acute nerve root disorders in these regions show no signs of classic ‘sympathetic abnormality’ when compared to patients with symptoms attributable to nerve injury beyond the root level, or from levels where roots contain sympathetic fibres. It has been suggested that the tortuous routes taken by sympathetic fibres may in itself present a greater potential for trauma and irritation (Pick 1970).
- As stated earlier, the cervical ganglia contain preganglionic fibres that may have their origins from nerve roots as low as T8.
This means that injuries or irritations of nerve roots in the upper and middle thoracic regions can have effects in the arms and head as well as in the viscera and somatic tissues more locally. Clinical investigations of upper extremity oedema, skin health, changes in circulation, changes in temperature or sweating must consider possible root origins in the thoracic spine.
- The major sympathetic vasoconstrictor supply to the upper limb arteries derives from roots T2 and T3.
The vasoconstrictor supply reaches the upper limb arteries via branches from the brachial plexus. This beautifully illustrates the complex anatomical routing of the sympathetic supply—from spinal cord, to ventral root of T2 and T3, into the ventral ramus, out via the white rami communicantes into the sympathetic chain—then synapsing with postganglionic fibres that course into the brachial plexus via grey rami communicantes before continuing on in the peripheral nerves to their vascular destinations. Changes in skin temperature and circulatory perfusion of the arm associated with musculoskeletal disorders like the chronic regional pain syndromes (CRPS I and II) may well relate to modest nerve injuries or irritations whose origins are in the upper thoracic regions.
Many postganglionic fibres that derive from the stellate ganglion supply the vasoconstrictor and sudomotor supply to the head; secretory fibres to the salivary glands; the pupil dilator muscles and muscles in the upper and lower eyelids.
Recall ‘Horners syndrome’ that results from destruction of this ganglion. Horner’s syndrome is characterised by constriction of the pupil, drooping of the upper eyelid (ptosis), enophthalmos (recession of the eyeball within the orbit) and absence of sweating on the face and neck. It is advisable for clinicians to seek out information relating to the face and eyes following significant neck traumas like whiplash.
Patients who have had simple tissue injuries, nerve injuries, spinal injuries or sprains/strains and some patients with symptoms classified as nerve root irritations quite often report symptoms and signs that are puzzling and not easy to explain. Knowledge of the complex innervation and workings of the sympathetic supply to the head, arms and legs can greatly help us, and often the patient too, to understand the possible origins of odd and understandably worrying symptoms. Common examples include: those related to temperature changes, changes in circulation, skin coloration, abnormal localised sweating, apparent and actual feelings of swelling, skin changes, visual disturbances, and dry mouth. Skilled clinicians may well ask about these types of symptoms in appropriate cases—and in so doing help to reassure the patient that what they describe is quite logical and reasonable when known anatomical features are taken into consideration.
Clinicians should also be on guard against an assumption of neural pathology when these types of symptoms are reported. For example it is well known that some patients with chronic pain syndromes demonstrate increased levels of somatic awareness. Patients are often anxious about their condition and their pain, more especially if they have been given an inadequate examination and explanation by those who the patient feels should know. The result can be that the patients’ attention becomes increasingly focused on sensations from their body, many which are either normal but are interpreted as abnormal or else relate to the physical manifestations of their distress. Clinicians are urged to be aware that symptoms of distress/anxiety often present in musculoskeletal tissues—for example many people when they are stressed get pain across the base of the neck and shoulders or in the low back.
In thoracic spinal injuries and presentations thought to have origins in the thoracic spine it is worth considering the SNS supply to the viscera and legs, and hence the potential for dysfunction in the relevant organs and tissues. Here again, enquiries about digestive, excretory and sexual function may be appropriate.
The sympathetic supply to the lower limb has its origins from T10, T11, T12 and L1–L2 occasionally L3 segments of the spinal cord. These spinal levels may be justifiably considered as possible sources in patients reporting ‘sympathetic’ effects in the lower limbs.
In all cases where the label ‘sympathetic’ symptoms is a possibility, great caution is advised in jumping to conclusions about pathology, pathological sources, symptom mechanisms and interventions. There is insufficient supportive evidence to pinpoint blame on the SNS unequivocally (see Chapters 2–5).
The effects of sympathetic and parasympathetic activity on target organs are summarised in Table 1.1.
The parasympathetic system
The traditional concept of the parasympathetic system is that it has an opposite and antagonistic effect to sympathetic activity on the organs and tissues innervated. However, Janig and Habler (1999) indicate that this state of affairs is the exception rather than the rule. In their overview of both systems (see Table 1.1) they state that:
- Most target tissues react only to one of the systems—the pacemaker cells of the heart are the exception.
- A few organs react to both—e.g., the iris, heart and urinary bladder.
- Most effects of both systems are excitatory, inhibition being rare.
Where there is reciprocal effect of the two systems on the target cells, it can usually be shown either that the systems work synergistically or that they exert their influence under different functional conditions. Two examples serve to illustrate this:
a) the opposite actions of sympathetic and parasympathetic systems on the size of the pupil is a consequence of the separate target cells supplied by each system – i.e. they go to different muscles;
b) fast changes of heart rate during changes of body position and emotional stress are generated via changes in activity in the parasympathetic neurons to the pacemaker cells; the sustained increase of heart rate during exercise is mainly generated by activation of sympathetic neurons supplying the heart.
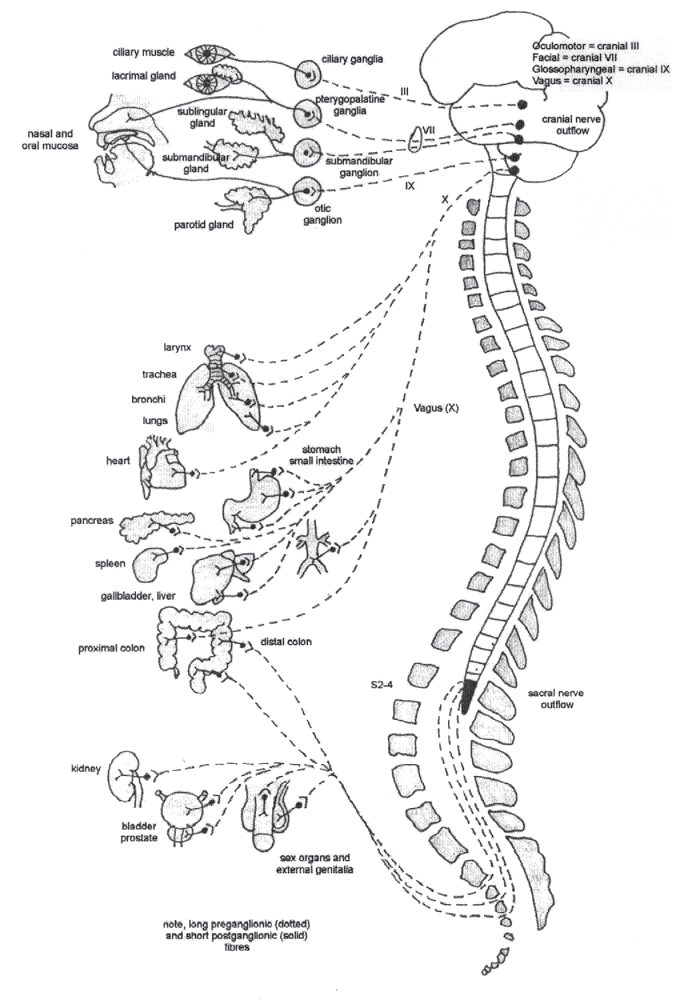
Because the parasympathetic system’s supply to its target tissues and organs is via four cranial nerves and nerves that have their origins in the second, third and fourth sacral nerves, the parasympathetic system is said to have a ‘craniosacral outflow’ (Figs 1.8, 1.5). Like the sympathetic system, the parasympathetic pathway consists of two neurones—a pre and postganglionic, with synapsing occurring between the two in parasympathetic ganglia. But unlike the sympathetic system, which mostly has short preganglionic and long postganglionic fibres, the parasympathetic system has very long preganglionic fibres. The postganglionic fibres tend to be short and derive from many very small ganglia that lie very close, or actually in, the target tissues. The ratio of pre to postganglionic fibres is between 1:15 and 1:20. Figure 1.8 summarises the supply.
Table 1.1 Effects of activation of sympathetic and parasympathetic neurones on autonomic target organs. (Adapted from Janig & Habler 1999)
| Organ and organ system | Activation of parasympathetic nerves | Activation of sympathetic nerves |
| Heart muscle | Decrease of heart rate Decrease of contractility (only atria) | Increase of heart rate Increase of contractility (atria, ventricles) |
| Blood vessels Arteries In skin of trunk and limbs In skin and mucosa of face In visceral domain In skeletal muscle In heart (coronary arteries) In erectile tissue In cranium Veins | 0 Vasodilation 0 0 Vasodilation Vasodilation (?) 0 | Vasoconstriction Vasoconstriction Vasoconstriction Vasoconstriction Vasodilation (cholinergic) Vasoconstriction Vasoconstriction Vasoconstriction Vasoconstriction Vasoconstriction Vasoconstriction |
| Gastrointestinal tract Longitudinal and circular muscle Sphincters | Increase of motility Relaxation | Decrease in motility Contraction |
| Capsule of spleen | 0 | Contraction |
| Urinary bladder Detrusor vesicae Trigone (internal sphincter) | Contraction 0 | Relaxation (small) Contraction |
| Reproductive organs Seminal vesicle, prostate Vas deferens Uterus | 0 0 0 | Contraction Contraction Contraction Relaxation — depends on species and hormonal state. |
| Eye Dilator muscle of pupil Sphincter muscle of pupil Ciliary muscle Tarsal muscle Orbital muscle | 0 Contraction (miosis) Contraction (accomodation) 0 0 | Contraction (mydriasis) 0 Contraction (lifting of lid) Contraction (protrusion of eye) |
| Tracheo-bronchial muscles | Contraction | Relaxation (probably mainly by adrenaline) |
| Piloerector muscles | 0 | Contraction |
| Exocrine glands Salivary glands Lachrymal glands Nasopharyngeal glands Bronchial glands Sweat glands Digestive glands (stomach, pancreas) Mucosa (small, large intestine) | Copious secretion Secretion Secretion Secretion 0 Secretion Secretion | Weak mucous secretion 0 ? Secretion (cholinergic) Decrease of secretion or 0 Decrease of secretion or reabsorption |
| Pineal gland | 0 | Increase in synthesis of melatonin |
| Brown and adipose tissue | 0 | Heat production |
| Metabolism Liver Fat cells β-cells in islets of pancreas | 0 0 Secretion | Glycogenolysis, glucongeogenesis Lipolysis (free fatty acids in blood increased) Decrease in secretion of pancreas |
| Adrenal medulla | 0 | Secretion of adrenaline and noradrenaline |
| Lymphoid tissue | 0 | Depression of activity (e.g. of natural killer cells) |
Oculomotor supply (cranial nerve III). The preganglionic fibres travel in the oculomotor nerve to the ciliary ganglia. Short postganglionic fibres supply the muscles of the eye and control lens focusing via the ciliary muscles.
Facial parasympathetic supply (cranial nerve VII).The facial nerve sends preganglionic fibres to two major ganglia—the pterygopalatine and the submandibular. The pterygopalatine postganglionic fibres supply glands in the lining of the mouth and nose and the tear glands. They also supply vasodilatory and secretory input to arteries, veins and glands of the face, nose, mouth, tongue, eyes and cerebrum. The postganglionic innervation deriving from the submandibular ganglia gives rise to a stimulatory supply to the submandibular and sublingual glands (salivary).
Glossopharyngeal parasympathetic supply (cranial nerve IX).Preganglionic fibres run to the otic ganglia that via its postganglionic fibres supplies the parotid gland. Some postganglionic neurons may also supply blood vessels of the jaw, the cerebral circulation and sweat glands around the lips.
Vagus parasympathetic supply (cranial nerve X).This dramatic nerve leaves the skull via the jugular foramen with the same dural sleeve as the glossopharyngeal and accessory nerves and descends into the abdomen. Its efferent parasympathetic supply is summarised in Figure 1.8.
Sacral parasympathetic outflow.Preganglionic fibres pass from the spinal cord segments S2–S4 to the ventral rami of the nerve roots. The fibres then pass within the roots down the spinal canal and out of their respective ventral foramen on the sacrum where they join to form the various branches and nerves of the sacral plexus. Parasympathetic supply leaves the sacral plexus as the pelvic splanchnic nerves. These nerves join the hypogastric plexus that derives from the pelvic part of the sympathetic chain. The sacral parasympathetic fibres supply the distal colon, rectum, bladder, prostate, kidney, sex organs and external genitalia. (Fig. 1.8)
Those readers who wish to know more about urogenital nerve supply and urogenital pain syndromes in men and women are advised to consult Wesselmann’s work (Wesselmann 1999, Wesselmann 2000).
Neurotransmitters and receptors of the ANS
The nervous system specialises in sending information—fast. Fast transmission of information involves impulses as a result of action potentials along the length of the axon. Where axons end they form synapses with other neurones or communicate with their target tissues via ‘neuroeffector’ junctions. The fast electrical message that arrives at the end of the axon produces its effect by stimulating the release of chemical messenger agents called neurotransmitters (see Fig. 1.6).
In the autonomic nervous system the peripheral pathway involves two neurones and therefore a single synapse between the dendrites of the preganglionic neurones and the ganglionic cell body of the postganglionic neurones (see Fig. 1.6). Preganglionic neurones of all sympathetic and parasympathetic neurones are said to be cholinergic because the neurotransmitter they liberate following the arrival of an action potential is acetylcholine. Acetylcholine then diffuses across the synaptic cleft and binds to receptor molecules on the cell membrane of the postganglionic cell body. This receptor binding process causes adjacent ion channels to open and a flow of ions to occur across the cell membrane. The resultant flow of ions across the cell membrane gives rise to a change in membrane voltage which in turn leads to the formation of a post synaptic action potential that then forms the impulse radiating to the nerve ends of the postganglionic fibre in the target tissues. The predominant acetylcholine receptor molecules in the postganglionic cell membrane are called nicotinic receptors, so named because the excitatory effect can be imitated by locally applied nicotine.
Adapted from: Harati Y, Machkhas H 1997 Spinal cord and peripheral nervous system. In: Low PA (ed) Clinical Autonomic Disorders 2nd edn. Lippincott-Raven, Philadelphia
The end terminals of the postganglionic nerve are often termed neuroeffector junctions. Here, the junctions of sympathetic and parasympathetic systems differ in the chemical transmitters they secrete.
All postganglionic neurones contain large numbers of varicosities (‘bags’ containing the neurotransmitters) strung along the axon branches where they are in contact with the target organ tissue. The chief sympathetic postganglionic neurone transmitter substance is norepinephrine (noradrenaline)—hence the sympathetic system in general is described as adrenergic. The parasympathetic neurone, on the other hand secretes acetylcholine—hence the term cholinergic.
In the case of the sympathetic system, noradrenaline is released in to the tissues and causes its effects there via noradrenaline receptors expressed on the tissues it supplies. These receptors are usually referred to as ‘adrenoreceptors’ and there are many ‘subtypes’ now known.
Adrenoreceptors may be found:
- On the cells of the tissues innervated.
- On any immune cells that may be present.
- On smooth muscle cells in local vasculature.
- On any sensory cells in the area, for example nociceptors.
- On the sympathetic terminals themselves—hence noradrenaline may be released by the sympathetic terminal and then act on it to produce further activity.
This ‘self stimulation’ is termed an autocrine effect. For example, there is now a good body of research demonstrating that released noradrenaline acts on adjacent adrenoreceptors on the postganglionic terminals to cause the production and release of prostaglandins that may cause nociceptor sensitisation or even pain (see Levine & Reichling 1999; see also Chapters 2–4).
An important principle for the clinician is to appreciate that any transmitter chemical like noradrenaline can only have an effect on a target tissue if active and relevant receptors for the chemical are present. Appreciate also, that chemical agents that activate the receptors are called ‘agonists’ and that ‘antagonists’ block the receptors and hence prevent their activity. Phentolamine, discussed in Chapter 4, is an example of an adrenoreceptor antagonist commonly used in the diagnosis and management of sympathetically maintained pain.
Readers interested in a more detailed account of the neurochemical organisation of the autonomic nervous system are advised to consult Milner et al (1999).
The enteric nervous system—the brain of the gut
The enteric nervous system is peculiar to the gastrointestinal tract and consists of a network of nerve cells and fibres embedded in its walls. It is considered to be a semiautonomous system with specific ‘programmes’ for motor responses, such as peristaltic reflexes and regional rate of contraction via its pacemaker systems (Camilleri 1997).
It consists of two major plexuses, the myenteric (Auerbach’s) and the submucous (Meissner’s) plexuses. There are also plexuses of nerve-fibre bundles in the muscle layers, mucosa, subserosa and around arteries. It is surprising to consider that there may be as many nerve cell bodies in the gut as there are in the spinal cord, about 100 million!! Further details can be found in Camilleri (1997).
According to Camilleri (1997), the current concept is that the enteric nerve complex has integrative sensory, interneurone and motor systems that can function independently of sympathetic and parasympathetic input from the CNS. It is believed that there are ‘hard-wired’ modules that detect the chemical and physical condition and contents of the gut, process the information in well established inter-neurone circuits and then mount appropriate motor responses involving control of secretory tissues and organs as well as muscular activity. Inputs from the CNS via the parasympathetic and sympathetic systems are thought to modulate the response patterns, and may even be able to ‘select’ particular responses to suit conditions elsewhere. CNS control is via the ‘excitatory’ parasympathetic vagal pathway on the one hand, and the sympathetic pathways that function to inactivate and inhibit gut muscle and digestive activities on the other.
The clinical message is that when under stress, threat or when life gets exciting, gut ‘activity’ and digestion is inhibited or even shut down—since it wastes vital energy that would otherwise be useful for survival. During stress our digestive secretions dry up (water conservation) and peristalsis halts. Diarrhoea associated with acute stress indicates however that one end of the system is active! From an evolutionary perspective (see Sapolsky 1994) this short lived event is a very adaptive strategy: first, clearing out the lower bowel (and bladder too) lightens the load to enable escape/more efficient action in the face of physical threat and second it makes you far less palatable to any predator due to the noxious smell and rather unsightly mess! The detrimental effects of ongoing stress are likely to include negative effects on bowel function—hence conditions like irritable bowel syndrome (reviewed in Chaitow 2000) that often occur concurrently in patients with chronic pain disability.
In times of low stress levels the gut is left unhindered to function efficiently. Patients with ongoing pain states, or who have pain states that are a source of high concern, are often highly distressed. The impact of this kind of ongoing and unresolved stress response on gut function and gut health is likely to be negative.
The visceral sensory system—the ‘autonomic or visceral afferents’
The viscera contains terminals of sensory neurones and very definitely has a sensory innervation (see e.g. Janig & Habler 1995). Visceral afferents may be mechanosensitive—being activated by various types of mechanical distortion (think bladder, gut, lungs and blood pressure) or chemically, for example picking up the osmolarity of the blood, or the concentrations of glucose. Some afferents may only be excited when the tissues are inflamed and therefore signal noxious events. Interestingly, vagal afferents innervating the liver may signal information in relation to toxins/infections picked up from the blood and as a result mediate general and protective sickness behaviours which includes a generalised increase in physical sensitivity (i.e. hyperalgesia) (Watkins et al 1995, Pennisi 1997, Sternberg & Gold 1997, Watkins 1997). It is certainly common to feel physically stiff and generally uncomfortable when you are suffering with an infection or are feeling unwell. Minor knocks and bumps can often be sickeningly painful. It seems a reasonable evolutionary tactic for the brain to put the somatic sensory systems on higher alert when we are in a more vulnerable state. Consider also that many patients in pain are well below ‘par’ and may already be nudged towards a hyperalgesic state. Add to this the toxic effects of medications and further shifts may occur.
Afferent fibres from the viscera travel to the central nervous system in the nerves and nerve plexuses described above for the sympathetic and parasympathetic systems. Thus, visceral afferents are found in the vagus and glossopharyngeal nerves and possibly other cranial nerves; also they are found in the second, third and fourth sacral nerves and in the thoracic and upper lumbar spinal nerves. Visceral sensory fibres can be divided into three types:
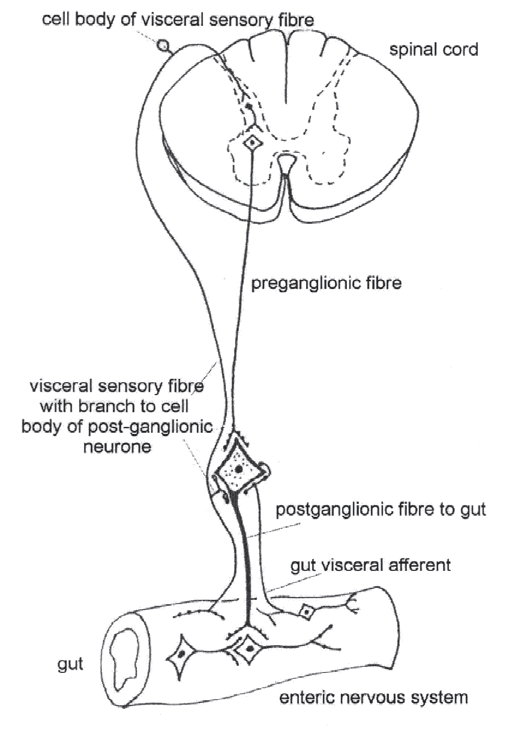
- Visceral afferents that project to the spinal cord (Fig. 1.9). Like their somatic counterparts these visceral sensory fibres have their cell bodies in the dorsal root ganglia and their central terminals in the outer lamina of the cord dorsal horn. These ‘spinal’ afferents travel from the viscera mainly in the splanchnic nerves and amount to only 1.5–2.0% of all spinal afferents that have their cell bodies in the dorsal root ganglia (Janig & Habler 1995). Clearly, the viscera has a relatively poor sensory supply when compared to the skin, where the supply is vast, or the deep somatic domain, where it is more modest than the skin, but still significant compared to the gut. As discussed below visceral primary afferents may send collateral branches to sympathetic postganglionic efferents in the prevertebral ganglia.
- Visceral afferents that travel to the CNS via cranial nerves and terminate in appropriate brain nuclei. For example, central endings of afferents travelling in the vagus nerve mainly terminate in the nucleus of the solitary tract (NST) that lies in the brain stem. Ritter et al (1992) state that 80–85% of nerve fibres in the vagus nerve are afferent fibres. Just like somatic sensory fibres, there are several functional subtypes of spinal and cranial visceral afferents. These are discussed in relation to visceral pain in the section following.
- ‘Enteric’ visceral afferents (Fig. 1.9). These afferents lie in the walls of the gut and communicate with enteric interneurones and motor neurones as well as sending branches to prevertebral ganglia where they synapse with postganglionic fibres of the autonomic system.
Visceral primary afferents continually sample the state of affairs in their target organs. Hence information about such things as the contents of the gut, bladder, colon, and the chemistry of the blood may be relayed centrally for processing and the generation of appropriate responses. Higher centres and ‘consciousness’ may become involved; we well know that situations in the viscera often promote physical action, such as when our bladder or bowel is full, when we are dehydrated, when we are hungry or satiated, and when sexually aroused. However, for many of the mundane operations that go on in the gut there is little need for conscious or subconscious analysis. Much activity and control goes on far nearer the tissues themselves.
Several ‘levels’ of integrative control have been identified:
- At the lowest integrative control level sits the enteric nervous system of the gut. Here, the neural organisation illustrates its isolated ‘sample — scrutinise — response’ capabilities and hence, its special autonomy.
- A second integrative level consists of a local control loop involving postganglionic autonomic fibres. Here, collateral branches of centrally projecting primary afferents as well as projections from visceral afferents of the enteric system (Fig. 1.9) synapse with postganglionic sympathetic fibres in the prevertebral ganglia. Hence, the formation of an ‘extraspinal’ reflex feedback loop. Regulation at this level is said to be via ‘extracentral reflexes’ since they can operate without involving the CNS, though they may well be influenced by modulating central inputs. These reflexes are thought to be important in control and regulation of gut motility (enhancement of peristalsis, storage function), regulation of fluid (excretion), as well as in protection. ‘Protection’ or protective reflexes that inhibit intestinal reflex activity might occur in relation to such things as inflammation of organs, peritonitis, overdistension or obstruction. Similar reflex pathways may also operate in the gall bladder and pancreas (Janig & Habler 1995).
- Visceral primary afferents are important for higher level integration involving reflex segmental spinal and cranial pathways as well as higher CNS processing and outputs.
Adapted from: Janig J, Habler, HJ 1995 Visceral-autonomic integration. In: Gebhart GF (ed) Visceral Pain. Progress in Pain Research and Management Vol. 5. IASP Press, Seattle
Effects of deep tissue pathology on somatic tissues and skin—can visceral disease precipitate musculosketal disorder?
In 1893 Henry Head (1893), an English neurologist, described how diseases of internal organs produced changes in skin areas that shared the same segmental origins. He described changes in sensitivity of the skin to touch, pressure and temperature that appeared in the acute phases of the disease and disappeared with recovery. For example, in pathology of the gall bladder, hyperalgesia can be found in skin derived from segments T6–T10. Later, Mackenzie (1909) observed hypertonic alterations and hypersensitivity in muscles belonging to the same segment as diseased organs. Some common sites of pain referral are reviewed in Figure 1.10. Proponents of connective tissue massage (CTM) have long considered the value of recognition and treatment of zones of abnormality in tension in the skin and subcutaneous layers relating to visceral dysfunction (reviewed in Luedecke 1969, Ebner 1975, Gifford J & Gifford L S 1988).
Clinical observations and research into disorders like the complex regional pain syndromes that have long been associated with ‘autonomic’ signs and symptoms document quite marked changes in ‘secondary’ tissues. Hence ‘referred’ signs and symptoms of pain, tenderness (hyperalgesia/ allodynia), altered tissue health, swelling, sweating, temperature, joint function, bone health and so forth.
It seems that lesions in nerves or pathology/dysfunction of visceral organs can lead to secondary symptoms and secondary physical changes, and hence pathology, in tissues that are related via innervation and connectivity. In support of this, there is even evidence that ‘visceral disease can actually be predicted from examination of trophic changes in skin and its appendages, subcutis, joint capsules, and fascia…with a probability of about 70%’ (Janig & Habler 1995.)
Multiple theories abound (see Janig & Habler 1995), most focusing round the concept that these secondary changes stem from the negative impact of ongoing activity in viscero-sympathetic reflex arcs. Research summarised by Janig and Habler (1995) demonstrates that increased visceral afferent activity from a cat’s bladder will cause a segmentally related increase in activity of sympathetic efferents involved in skin sweating and muscle circulation and a decrease in activity in sympathetic efferents responsible for skin circulation. Thus, noxious input due to excessive distension of the bladder, or experimental inflammation of it, causes increased activity in all visceral afferent fibre types. This includes nociceptors. This activity bears a direct relation to activity in the segmentally related sympathetic efferent fibres that control sweating and circulation. The findings show that noxious input produces a significant decrease in circulation to muscles, an increase in sweating and an increase in circulation to the skin. The research also demonstrates significant ongoing activity for several hours after the provocation abates. It is not difficult to envisage that ongoing activity of days or weeks could lead to prolonged changes in circulation leading to trophic and other changes in the tissues affected. For example, prolonged increases in circulation to skin could result in oedema and prolonged circulatory deprivation to muscle could cause muscle fibre degeneration, dysfunction and impairment.
The disciples of a holistic approach to pain and health have been around for a long time; the evidence that they are right is only now emerging. One system’s health or ill-health appears to have the potential to significantly impact traditionally unrelated tissues.
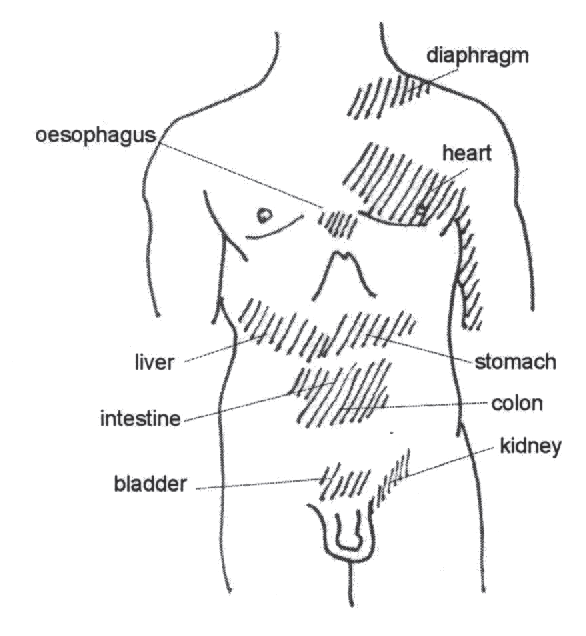
Adapted from: Westmorland et al 1994 Medical Neurosciences 3rd edn. Little Brown, Boston
The autonomic innervation of the immune system, stress and mind-body links
One area of recent interest to mind body pathways and the science of psychoneuroimmunology is the finding that tissues where immune cells develop have a rich autonomic innervation (Watkins 1997). This innervation has been given little attention by pain researchers, yet it may be of considerable relevance.
Thus, sympathetic and parasympathetic fibres are found in bone marrow, the thymus gland and lymphoid tissue and organs generally. It seems that the activity of both branches of the autonomic nervous system have the capacity to modulate the maturity and activation of the immune cells, especially in those areas like the airways and gut that are exposed to antigens and allergens. Watkins (1997) states: ‘Although the exact role of autonomic innervation in regulating immunity has not been clearly defined, there is sufficient evidence to suggest that autonomic nerves are capable of regulating almost all the cells involved in inflammation.’
There is some evidence to suggest that age-related dysfunction of the ANS, including that to the lymphoid tissue, may be responsible for the agerelated decay in immunological function (see references in Watkins 1997 p. 17). One message is that we should look after our ANS for as long as we can!
Since the activity of the ANS is powerfully influenced by perception of our circumstances, by emotions and by our mood state, this physical linking to the immune system holds interest to those who embrace the need for multidimensional models of health and disease.
The scientific scrutiny of mind-body links and pathways is gathering a great deal of useful and interesting information for the multidimensionally comfortable clinician. For example, in discussing the evidence for cortical control of immunity Watkins (1997 p.17) reports:
- That the effects of stress, perception and personality on immunity have all been extensively investigated (see below.)
- That there appears to be a relationship between cerebral dominance and allergy and auto-immune disease—with left handers having a 11.5-fold increase in incidence of self-reported allergy.
- Lesions of the cortex produced by electrolytic ablation have shown that immunity is suppressed with lesions to the left cortex and enhanced with ablations to the right!
Stress is known to activate central nervous system pathways that mediate the activity of the sympathetic adreno–medullary (SAM) axis—hence increasing levels of circulating adrenaline, as well as the hypothalamic– pituitary–adreno-cortex (HPA) axis that results in increased levels of the circulating steroid cortisol. It has long been postulated that ongoing stress, and hence the effects mediated by these hormones or via the direct pathways detailed above, may have detrimental effects and even produce or promote somatic disease processes as well as changes in mood (e.g. Selye & Tuchweber 1976, Selye 1978, Sapolsky 1994, Martin 1997, Watkins 1997). While fluctuations in stress are a normal part of everyday life and something that evolution has catered for, ongoing stress with no let up is not.
Some points
Stressful situations activate the SAM axis resulting in the fight or flight response and associated feelings of anger and anxiety.
In contrast, activation of the HPA axis promotes submission, and the emotions of defeat and despair. Typically, chronically stressful situations promote a vacillation between anger and despair as individuals fight to gain control over their situation or give up, believing they have no control. ‘These cycles of anger and despair promote the production of a destructive range of catabolic hormones, injurious to a number of bodily systems, not just the immune system’ (see Watkins 1997, p11).
Animal studies have demonstrated immunosuppresive effects of a wide range of stressors, for example, isolation, separation, overcrowding, introduction of an aggressive intruder, restraint, cold, noise and inescapable foot shock (see Watkins 1997). It seems relatively easy to see the possible links to patients who suffer with ongoing pain syndromes.
Chronic stress has been shown to impair wound healing in humans. Death of a spouse suppresses the immune system for 2–6 weeks. Traumatic marital separation has been shown to be even more immunosuppresive than bereavement. However, Watkins (1997), states that ‘current opinion now believes that minor chronic stress, termed “microstress,” is more immunosuppresive than single life events such as bereavement.’
In contrast, animal studies have shown increased survival of animals exposed to influenza virus who also underwent restraint stress. The way a stressor is perceived, or the ‘will to survive’ may have a huge impact on whether there is a positive or negative impact on the immune system or other potentially destructive processes that stress can precipitate. Clinically, the key may be to improve patients’ ability to cope, and increase their feelings of control and of self-esteem. If patients are unable to learn the skills that are needed to pick themselves up, and if they continue to see their situation as hopeless, then successful outcomes are unlikely. It seems that the ‘will to survive’ or at least a positive belief in one’s ability to cope, get control, or overcome a situation may have a significant multidimensional health promoting impact.
With response to stress, there is a great inter-individual variability and a marked spectrum to consider. What scares you or makes you feel cautious may not necessarily have the same effect on the next person. But also, what scares you now may not scare you in the future, and what doesn’t bother you now, may well do so in years to come. Past experiences, our inborn temperament and our learned and instinctive coping strategies may all have a part to play in how we cope right now.
There is some evidence that psychosocial disruption in utero, postnatally, and during childhood can have long-term consequences on the immune system. Early emotionally traumatic years may manifest in poorer coping responses to stress in adulthood. Psychological factors, such as our coping strategies, have been shown to be as important as the characteristics of the stress itself, in determining the immunological consequences of stress and the outcome of disease. For example, coping strategies and psychological factors have been shown to be significant predictors of who dies from acute asthma, in addition to predicting the progression of viral infections, AIDS, cancer and heart disease. On a positive note is the evidence that some coping strategies can minimise the detrimental effects of stressful life events; that positive emotional states can enhance immunity (for all references see Watkins 1997), and that some people’s poor coping strategies can be shifted into more helpful and health promoting ones. Clinicians are urged to consider the potentially massive positive psychophysiological impacts that can be achieved as a result of treatment encounters, and that the ANS is regarded as a major efferent pathway linking mind and body.
REFERENCES AND FURTHER READING
Camilleri M 1997 Autonomic regulation of gastrointestinal motility. In: Low P A (ed) Clinical Autonomic Disorders. Lippincott-Raven, Philiadelphia 135–145
Chaitow L 2000 Fibromyalgia Syndrome. A Practitioners guide to treatment. Churchill Livingstone, Edinburgh
Damasio AR 1995 Descartes’ Error: Emotion, reason and the human brain. Picador, London
Damasio AR 2000 The feeling of what happens. Body, emotion and the making of consciousness. Vintage, London
Ebner M 1975 Connective Tissue Massage, Theory and Therapeutic application. R E Kreiger, New York
Gifford LS 1998 The mature organism model. In: Gifford LS (ed) Topical Issues in Pain 1. Whiplash—science and management. Fear-avoidance beliefs and behaviour. CNS Press, Falmouth 45–56
Gifford LS 1998a Central mechanisms. In: Gifford LS (ed) Topical Issues in Pain 1. Whiplash—science and management. Fear-avoidance beliefs and behaviour. CNS Press, Falmouth 67–80
Gifford LS 1998b Output mechanisms. In: Gifford LS (ed) Topical Issues in Pain 1. Whiplash—science and management. Fear-avoidance beliefs and behaviour. CNS Press, Falmouth 81–91
Gifford LS (ed) 2002 Topical Issues in Pain 4. Placebo. Muscles and Pain. Pain Management. CNS Press, Falmouth
Gifford J, Gifford LS 1988 Connective tissue massage. In: Wells PE, Frampton V, Bowsher D (eds) Pain Management by Physiotherapy. Butterworth Heinemann, Oxford 213–227
Grieve G P 1994 The autonomic nervous system in vertebral pain syndromes. In: Boyling JD, Palastanga N (eds) Grieve’s Modern Manual Therapy. Churchill Livingstone, Edinburgh 293–308
Harati Y, Machkhas H 1997 Spinal cord and peripheral nervous system. In: Low PA (ed) Clinical Autonomic Disorders 2nd edn. Lippincott-Raven, Philadelphia 28
Head H 1893 On disturbances of sensation with especial reference to the pain of visceral disease. Brain 16: 1–132
Janig W, Habler H-J 1995 Visceral-autonomic integration. In: Gebhart GF (ed) Visceral Pain. Progress in Pain Research and Management Vol 5. IASP Press, Seattle 311–348
Janig W, Habler H-J 1999 Organization of the autonomic nervous system: structure and function. In: Appenzeller O (ed) Handbook of Clinical Neurology Vol 74 (30): The Autonomic Nervous System. Part 1. Normal Functions. Elsevier,
Amsterdam 1–52
LeDoux J 1998 The Emotional Brain. The mysterious underpinnings of emotional life. Weidenfeld & Nicolson, London
Levine JD, Reichling DB 1999 Peripheral mechanisms of inflammatory pain. In: Wall PD, Melzack R (eds) Textbook of Pain 4th edn. Churchill Livingstone, Edinburgh 59–84
Luedecke U 1969 History, basis and technique of connective tissue massage. Australian Journal of Physiotherapy 15(4): 141–148
Mackenzie J 1909 Symptoms and their Interpretation. Shaw and Sons, London
Martin P 1997 The Sickening Mind. Brain, behaviour, immunity and disease. Harper Collins, London
Meyers GE 1986 William James: His life and thought. Yale University Press, New Haven, Connecticut
Milner P, Lincoln J, Burnstock G 1999 The neurochemical organisation of the autonomic nervous system. In: Appenzeller O (ed) Handbook of Clinical Neurology Vol 74 (30): The Autonomic Nervous System. Part 1. Normal Functions. Elsevier, Amsterdam 87–133
Pick JP (1970) The Autonomic Nervous System: Morphological, Comparative, Clinical and Surgical Aspects. Lippencott. Philadelphia
Pennisi E 1997 Tracing molecules that make the brain-body connection. Science 275: 930–931
Ritter S, Ritter R C, Barnes C D (eds) 1992 Neuroanatomy and Physiology of Abdominal Vagal Afferents. CRC Press, Boca Raton, Fla.
Sapolsky R M 1994 Why Zebras don’t get ulcers. A guide to stress, stress-related diseases, and coping. Freeman, New York
Selye H 1978 The Stress of Life. McGraw Hill, New York Selye H, Tuchweber B 1976 Stress in relation to aging and disease. In: Everitt A,
Burgess J (eds) Hypothalamus, Pituitary and Aging. Charles C Thomas, Springfield
Sternberg EM, Gold PW 1997 The mind-body interaction in disease. Scientific American Special Issue: Mysteries of the Mind June: 8–15
Thacker MA 1998 Whiplash—is there a lesion? In: Gifford LS (ed) Topical Issues in Pain 1. Whiplash—science and management. Fear-avoidance beliefs and behaviour. CNS Press, Falmouth 27–43
Watkins A (ed) 1997 Mind-body Medicine. A clinicians guide to psychoneuroimmunology. Churchill Livingstone, Edinburgh
Watkins A 1997 Mind-body pathways. In: Watkins A (ed) Mind-body Medicine. A clinicians guide to psychoneuroimmunology. Churchill Livingstone, Edinburgh 1–25
Watkins LR, Maier SF, Goehler LE 1995 Immune activation: the role of proinflammatory cytokines in inflammation, illness responses and pathological pain states. Pain 63: 289–302
Wesselmann U 1999 Pain of pelvic origin (urological and gynecological). In: Max M (ed) Pain 1999—an updated review. Refresher course syllabus. IASP Press, Seattle 47–58
Wesselmann U 2000 Urogenital pain syndromes in men and women. In: Devor M, Rowbotham MC, Wiesenfeld-Hallin Z (eds) Proceedings of the 9th World Congress on Pain. Progress in Pain Research and Management, Vol 16 IASP
Press, Seattle 551–566
Westmoreland BF, Benarroch EE, Daube JR et al 1994 Medical Neurosciences. Little Brown, Boston
Williams PL, et al (eds) 1995 Gray’s Anatomy 38th edn. Churchill Livingstone, Edinburgh